Биология в лицее
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ 







Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Транскрипция и трансляция
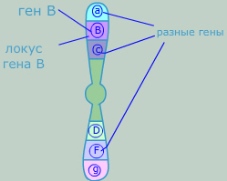 Ген (греч. génesis — происхождение) — это элементарная единица наследственности, представляющая отрезок молекулы ДНК (у некоторых вирусов РНК).
Ген (греч. génesis — происхождение) — это элементарная единица наследственности, представляющая отрезок молекулы ДНК (у некоторых вирусов РНК).
Существование дискретных наследственных факторов предположил Грегор Мендель в 1865 году, а в 1909 году В. Иогансен назвал их генами.
Ген — участок молекулы ДНК, содержащий информацию о первичной структуре одной полипептидной цепочки или молекулы рРНК и тРНК. Таким образом, ген определяет строение одного из белков живой клетки и тем самым участвует в формировании признака или свойства организма.
Матричная и кодирующая цепи ДНК
Две цепи ДНК в области гена принципиально различаются по своей функциональной роли: одна из них является кодирующей , или смысловой, вторая, комплементарная кодирующей цепи, — матричной .
Это значит, что в процессе «считывания» гена (транскрипции, или синтеза мРНК) в качестве матрицы выступает только одна — матричная — цепь ДНК. Продукт же этого процесса — мРНК — по последовательности нуклеотидов совпадает с кодирующей цепью ДНК (с заменой тиминовых оснований на урациловые).
Таким образом, получается что с помощью матричной цепи ДНК при транскрипции воспроизводится в структуре РНК генетическая информация кодирующей цепи ДНК.
Информация на кодирующей цепи записана в направлении от 5‘-конца к 3‘-концу. И этот же конец принято считать 5′-концом всего гена (хотя у его матричной цепи здесь находится 3′-конец).
Всего на длинной молекуле ДНК находится несколько тысяч генов. И, как правило, для всех этих генов кодирующей является одна и та же цепь ДНК. Но иногда бывает иначе: для одних генов в качестве смысловой выступает одна цепь ДНК, а для других генов — противоположная цепь. Такие гены, очевидно, читаются в разных направлениях. Подобная ситуация обнаружена, в частности, некоторых генов у дрозофилы.
Значение биосинтеза белка в процессах жизнедеятельности
Каждая живая клетка создаёт вещества, образующие её организм. Этот процесс называют биосинтезом. Реакции, обеспечивающие этот процесс, ферментативные, связаны с потреблением энергии и функцией внутриклеточных структур. Например, синтез углеводов в растительной клетке связан с энергией света и хлоропластами, а биосинтез белка — с энергией химических связей АТФ и рибосомами.
В биосинтезе молекул белка участвуют 20 видов аминокислот, разные виды РНК и многочисленные рибосомы, расположенные на мембранах ЭПС.
Биосинтез протекает в течение всей жизни клетки, но для каждого вида ткани характерны специфические белки. В костной ткани это оссеин, в мышечной — актин и миозин, в крови фибриноген, антитела и др. Каждый белок имеет свой состав аминокислот и последовательность их соединения, что определяет функции и свойства белковой молекулы.
Для продолжения изучения темы Вам необходимо вспомнить особенности строения молекулы ДНК.
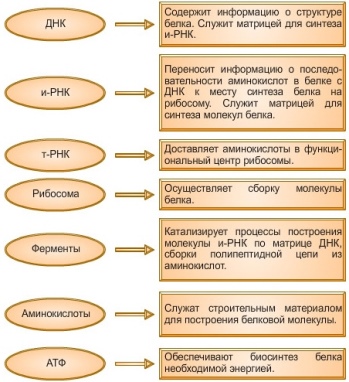
ДНК служит матрицей для синтеза иРНК, которая переносит наследственную информацию из ядра к рибосоме, месту синтеза полипептидной цепи.
Возникшая цепочка иРНК через некоторое время отделяется от ДНК, направляется через ядерные поры в каналы ЭПС и доставляется на рибосомы. Синтез иРНК происходит на одной из нитей ДНК, точка начала синтеза — промотор , точка окончания — терминатор .

Процесс синтеза иРНК называют транскрипция . иРНК — это продукт, содержащий копии гена или группы генов.
Транскрипция
Структура любой белковой молекулы закодирована в ядерной ДНК, которая непосредственного участия в её синтезе не принимает. Она служит лишь матрицей для синтеза информационной РНК (иРНК), которая является переносчиком наследственной информации из ядра к рибосоме, месту синтеза полипептидной нити белка.
Процесс биосинтеза белка включает в себя ряд последовательно протекающих событий.
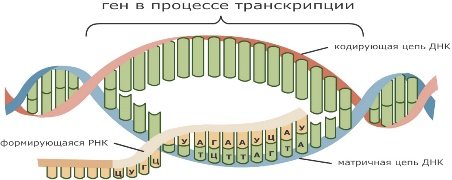
При биосинтезе часть молекулы ДНК расщепляется на две цепочки, на одной из которых синтезируется иРНК. Она представляет собой негативную фотографию последовательности нуклеотидов ДНК. Этот процесс считывания информации обеспечивается работой фермента — полимераза , которая, двигаясь по расплетённому участку ДНК, подбирает с учётом комплементарности нуклеотиды и соединяет их. Эта иРНК содержит полную информацию о порядке расположения аминокислот в белке.
Роль тРНК в биосинтезе белка. Трансляция
К месту сборки белка поступают аминокислоты. Сюда их доставляют тРНК. Эти молекулы имеют форму клеверного листа, на вершине которого находятся три нуклеотида — триплет, или антикодон, кодирующий определённую аминокислоту.
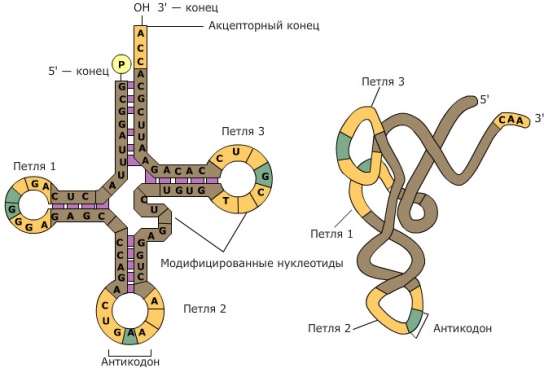
Каждая тРНК может соединиться лишь с одной молекулой аминокислоты и доставить её к рибосоме. Например: антикодон в тРНК АЦГ комплементарен триплету УГЦ в иРНК. Эти два триплета кодируют аминокислоту серин. В ДНК серину будет соответствовать триплет АЦГ. Сравнение триплетов ДНК и тРНК показывает, что они одинаковы.
Активацию определённой аминокислоты осуществляет свой особый фермент. Механизм активации заключается в том, что фермент одновременно взаимодействует с аминокислотой и с АТФ, которая при этом теряет пирофосфат. Образуется тройной комплекс из фермента, аминокислоты и тРНК, способный сразу образовывать пептидную связь. Без такого взаимодействия свободная аминокислота не может образовывать пептидную связь.
Трансляция (от лат. translatio — «передача») — процесс синтеза полипептидных цепей на матрице информационной РНК в рибосомах.
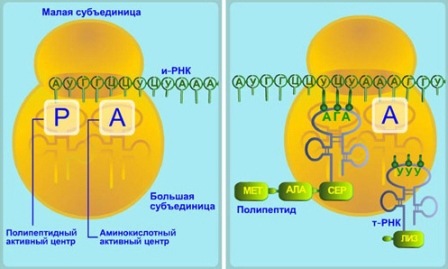 На иРНК нанизывается рибосома, и начинается синтез белка. В двух активных центрах рибосомы — А — аминокислотном и Р — пептидном — помещается всего шесть нуклеотидов иРНК (два триплета). К центру А из цитоплазмы всё время подходят разные тРНК с аминокислотами. Здесь происходит узнавание аминокислот. Если антикодон тРНК и кодон иРНК оказываются комплементарными друг другу, то тРНК вместе с аминокислотой переплывает с помощью ферментов в центр Р, в котором происходит освобождение тРНК от аминокислоты. Между последней аминокислотой, уже синтезированной части белка, и вновь доставленной аминокислотой возникает пептидная связь.
На иРНК нанизывается рибосома, и начинается синтез белка. В двух активных центрах рибосомы — А — аминокислотном и Р — пептидном — помещается всего шесть нуклеотидов иРНК (два триплета). К центру А из цитоплазмы всё время подходят разные тРНК с аминокислотами. Здесь происходит узнавание аминокислот. Если антикодон тРНК и кодон иРНК оказываются комплементарными друг другу, то тРНК вместе с аминокислотой переплывает с помощью ферментов в центр Р, в котором происходит освобождение тРНК от аминокислоты. Между последней аминокислотой, уже синтезированной части белка, и вновь доставленной аминокислотой возникает пептидная связь.
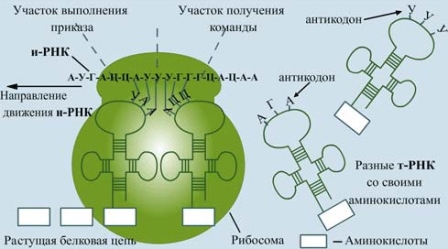 Молекула тРНК отдаёт аминокислоту и вновь перемещается в цитоплазму, где снова присоединяет такую же аминокислоту, а рибосома перемещается на один триплет влево. Постепенно за счёт присоединения аминокислот нить белка удлиняется.
Молекула тРНК отдаёт аминокислоту и вновь перемещается в цитоплазму, где снова присоединяет такую же аминокислоту, а рибосома перемещается на один триплет влево. Постепенно за счёт присоединения аминокислот нить белка удлиняется.
Если антикодон тРНК и кодон иРНК не комплементарны, то тРНК с аминокислотой уходят к другим иРНК и рибосомам.
Пептидная цепочка удлиняется до тех пор, пока не закончится трансляция и рибосома не соскочит с иРНК. Полипептидная цепочка погружается в канал ЭПС и там приобретает вторичную, третичную или четвертичную структуры.
Скорость сборки одной молекулы белка, состоящего из 200 — 300 аминокислот, составляет 1 — 2 минуты.
В процессе биосинтеза белка реализуются функции многих веществ и органоидов клетки и используется энергия АТФ.
Регуляция биосинтеза белка
 Работа генов в любом организме — прокариотическом или эукариотическом — контролируется и координируется.
Работа генов в любом организме — прокариотическом или эукариотическом — контролируется и координируется.
Различные гены обладают неодинаковой временной активностью. Одни из них характеризуются постоянной активностью. Это гены, которые отвечают за синтез белков, необходимых или организму на протяжении всей жизни, например ферменты.
Большинство генов обладает непостоянной активностью, они синтезируются, когда это необходимо клетке.
Различают структурные и регуляторные белки клетки. Структурные выполняют ферментативную, транспортную и структурную функции, а регуляторные управляют синтезом структурных генов.
Регуляция синтеза белка осуществляется на всех его этапах. Регуляция происходит на генетическом уровне. Есть сложный механизм «включения» и «выключения» генов на разных этапах жизни клетки на уровне оперона .
Оперон — совокупность генов, которые расположены рядом на ДНК, контролируют один процесс и регулируются одними и теми же элементами.
Оперон является функциональной единицей транскрипции у прокариот.
В состав оперона прокариот входят структурные гены и регуляторные элементы. Структурные гены кодируют белки, осуществляющие последовательно этапы биосинтеза какого-либо вещества. Их может быть один или несколько, в ходе транскрипции они работают как единый ген. На них синтезируется единая молекула иРНК.
![]() Регуляторные элементы
Регуляторные элементы
Промотор — это место начала транскрипции. Оно представлено нуклеотидами ДНК, с которыми связывается белок-фермент РНК-полимераза. Промотор определяет, какая из двух цепей будет служить матрицей для и-РНК.
Оператор — участок связывания регуляторного белка с ДНК. Этот белок репрессор, т.е. подавитель или белок-активатор процесса транскрипции. Оператор — это начало считывания генетической информации. На работу этих белков влияют вещества субстраты, которые могут взаимодействовать с белком-репрессором, освобождая проход для полимеразы, обеспечивая этим синтез и-РНК. Пока репрессор находится на операторе, полимераза не может сдвинуться с места и начать синтез.
Терминатор — участок в конце оперона, сигнализирующий о прекращении транскрипции.
Регуляция генной активности у эукариот сложнее, чем у бактерий.
В отличие от прокариот, образующиеся в ядре иРНК подвергаются ряду изменений. Сначала синтезируется длинная иРНК, а затем ферменты вырезают из неё участки, не несущие информацию о строении белка.
У эукариот, наряду с регуляторными процессами, влияющими на функции отдельной клетки, существуют системы регуляции целого организма. Гормоны образуются в клетках желёз внутренней секреции и с кровью разносятся по всему телу; регулируют процессы синтеза иРНК и белков только в клетках-мишенях. Гормоны связываются с белками-рецепторами клеточных мембран и включают системы изменения структуры клеточных белков, которые влияют на синтез белков на рибосомах и на транскрипцию определённых генов. Так, адреналин включает синтез ферментов, расщепляющих гликоген мышц до глюкозы, а инсулин способствует синтезу гликогена из глюкозы в печени.
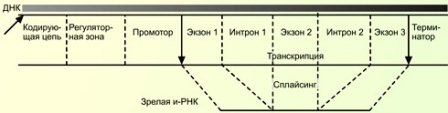 Особенность эукариотических генов — прерывистость. Это означает, что область гена, кодирующего белок, состоит из нуклеотидных последовательностей двух типов экзонов и интронов . Экзон — это участок ДНК, несущий информацию о структуре белка, Интрон — некодирующая область гена. Во время траскрипции сначала образуется иРНК, несущая информацию об экзонах и интронах, а затем происходит вырезание интронов и сшивание оставшихся экзонов ферментами. Этот процесс называется сплайсинг . В итоге образуется иРНК, которая служит матрицей для трансляции.
Особенность эукариотических генов — прерывистость. Это означает, что область гена, кодирующего белок, состоит из нуклеотидных последовательностей двух типов экзонов и интронов . Экзон — это участок ДНК, несущий информацию о структуре белка, Интрон — некодирующая область гена. Во время траскрипции сначала образуется иРНК, несущая информацию об экзонах и интронах, а затем происходит вырезание интронов и сшивание оставшихся экзонов ферментами. Этот процесс называется сплайсинг . В итоге образуется иРНК, которая служит матрицей для трансляции.
Ген эукариот похож на оперон прокариот, хотя и отличается более протяжённой регуляторной зоной и тем, что он кодирует один белок, а не несколько, как оперон у бактерий. Познание регуляторных механизмов транскрипции и трансляции необходимо для управления процессами реализации генетической информации.
Источник
Смысл (молекулярная биология) — Sense (molecular biology)
В области молекулярной биологии и генетики , то смысл из нуклеиновой кислоты молекулы, в частности , в виде нити ДНК или РНК , относится к характеру роли нити и ее комплементом в указании последовательности аминокислот . В зависимости от контекста смысл может иметь несколько разные значения. Например, отрицательно-смысловая цепь ДНК эквивалентна матричной цепи, тогда как положительно-смысловая цепь — нематричная цепь, нуклеотидная последовательность которой эквивалентна последовательности транскрипта мРНК.
СОДЕРЖАНИЕ
Чувство ДНК
Из-за комплементарного характера образования пар оснований между полимерами нуклеиновых кислот двухцепочечная молекула ДНК будет состоять из двух цепей с последовательностями, которые являются обратными комплементами друг друга. Чтобы помочь молекулярным биологам конкретно идентифицировать каждую нить индивидуально, две нити обычно различают как «смысловую» цепь и «антисмысловую» цепь. Отдельная цепь ДНК называется положительно-смысловой (также положительной (+) или просто смысловой ), если ее нуклеотидная последовательность напрямую соответствует последовательности транскрипта РНК, который транслируется или переводится в последовательность аминокислот (при условии, что любая основания тимина в последовательности ДНК заменены основаниями урацила в последовательности РНК). Другая цепь двухцепочечной молекулы ДНК называется отрицательно-смысловой (также отрицательной (-) или антисмысловой ) и является обратно комплементарной как положительно-смысловой цепи, так и транскрипту РНК. На самом деле именно антисмысловая цепь используется в качестве матрицы, из которой РНК-полимеразы конструируют транскрипт РНК, но комплементарное спаривание оснований, с помощью которого происходит полимеризация нуклеиновой кислоты, означает, что последовательность транскрипта РНК будет выглядеть идентичной смысловой цепи, за исключением от использования транскрипта РНК урацила вместо тимина.
Иногда вместо смыслового и антисмыслового словосочетания « кодирующая цепь» и « матричная цепь» встречаются, соответственно, и в контексте двухцепочечной молекулы ДНК использование этих терминов по существу эквивалентно. Однако кодирующая / смысловая цепь не всегда должна содержать код, который используется для создания белка; могут транскрибироваться как кодирующие белок, так и некодирующие РНК .
Термины «смысл» и «антисмысловой» относятся только к конкретному рассматриваемому транскрипту РНК, а не к цепи ДНК в целом. Другими словами, любая цепь ДНК может служить смысловой или антисмысловой цепью. Большинство организмов с достаточно большими геномами используют обе цепи, каждая из которых функционирует как матричная цепь для разных транскриптов РНК в разных местах одной и той же молекулы ДНК. В некоторых случаях транскрипты РНК могут транскрибироваться в обоих направлениях (т.е. на любой цепи) из общей промоторной области или транскрибироваться из интронов на любой цепи (см. «Амбисенс» ниже).
Антисмысловая ДНК
Смысловая цепь ДНК выглядит как транскрипт информационной РНК (мРНК) и поэтому может использоваться для считывания ожидаемой кодоновой последовательности, которая в конечном итоге будет использоваться во время трансляции (синтеза белка) для построения аминокислотной последовательности, а затем белка. Например, последовательность «ATG» в смысловой цепи ДНК соответствует кодону «AUG» в мРНК, который кодирует аминокислоту метионин . Однако сама смысловая цепь ДНК не используется в качестве матрицы для мРНК; именно антисмысловая цепь ДНК служит источником белкового кода, поскольку с основаниями, комплементарными смысловой цепи ДНК, она используется в качестве матрицы для мРНК. Поскольку в результате транскрипции образуется продукт РНК, комплементарный цепи матрицы ДНК, мРНК комплементарна антисмысловой цепи ДНК.
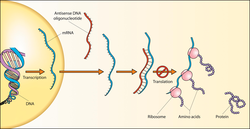
Следовательно, триплет оснований 3′-TAC-5 ‘в антисмысловой цепи ДНК (комплементарный 5′-ATG-3′ смысловой цепи ДНК) используется в качестве матрицы, что приводит к 5’-AUG-3 ‘ триплет оснований в мРНК. Смысловая цепь ДНК будет иметь триплет ATG, который похож на триплет мРНК AUG, но не будет использоваться для производства метионина, потому что он не будет использоваться напрямую для создания мРНК. Смысловая цепь ДНК называется «смысловой» цепью не потому, что она будет использоваться для производства белка (это не будет), а потому, что она имеет последовательность, которая непосредственно соответствует последовательности кодона РНК. По этой логике сам транскрипт РНК иногда называют «смысловым».
Пример с двухцепочечной ДНК
Некоторые области внутри двухцепочечной молекулы ДНК кодируют гены , которые обычно представляют собой инструкции, определяющие порядок, в котором аминокислоты собираются для образования белков, а также регуляторные последовательности, сайты сплайсинга , некодирующие интроны и другие генные продукты . Чтобы клетка могла использовать эту информацию, одна цепь ДНК служит шаблоном для синтеза комплементарной цепи РНК . Транскрибируемая цепь ДНК называется цепочкой-матрицей с антисмысловой последовательностью, а полученный из нее транскрипт мРНК называется смысловой последовательностью (комплемент антисмысловой последовательности). Также говорят, что нетранскрибируемая цепь ДНК, комплементарная транскрибированной цепи, имеет смысловую последовательность; он имеет ту же смысловую последовательность, что и транскрипт мРНК (хотя Т-основания в ДНК заменены на U-основания в РНК).
| 3′CGCTATAGCGTTT 5 ′ | Антисмысловая цепь ДНК (матричная / некодирующая) | Используется как шаблон для транскрипции. |
| 5′GCGATATCGCAAA 3 ′ | Смысловая цепь ДНК (не шаблонная / кодирующая) | Дополняет прядь шаблона. |
| 5′GCGAUAUCGCAAA 3 ′ | смысловой транскрипт мРНК | Нить РНК, которая транскрибируется с некодирующей (матричной / антисмысловой) цепи. Примечание 1. За исключением того факта, что все тимины теперь являются урацилами ( T → U ), они комплементарны некодирующей (матричной / антисмысловой) цепи ДНК и идентичны кодирующей (не шаблонной / смысловой) цепи ДНК. |
| 3′CGCUAUAGCGUUU 5 ′ | антисмысловой транскрипт мРНК | Нить РНК, которая транскрибируется с кодирующей (не шаблонной / смысловой) цепи. Примечание. За исключением того факта, что все тимины теперь являются урацилами ( T → U ), они комплементарны кодирующей (не шаблонной / смысловой) цепи ДНК и идентичны некодирующей (матричной / антисмысловой) цепи ДНК. |
Имена, присвоенные каждой нити, фактически зависят от того, в каком направлении вы пишете последовательность, содержащую информацию для белков («смысловую» информацию), а не от того, какая нить изображена как «вверху» или «внизу» (что произвольно). Единственная биологическая информация, которая важна для маркировки цепей, — это относительное расположение концевой 5′-фосфатной группы и концевой 3′-гидроксильной группы (на концах рассматриваемой цепи или последовательности), поскольку эти концы определяют направление транскрипции и перевод. Последовательность, записанная 5′-CGCTAT-3 ‘, эквивалентна последовательности, записанной 3′-TATCGC-5’, если отмечены концы 5 ‘и 3’. Если концы не помечены, предполагается, что обе последовательности написаны в направлении от 5′-к-3 ‘. «Нить Ватсона» относится к верхней нити от 5’-к-3′(5 ‘→ 3′), тогда как «Криковская цепь» относится к нижней нити от 5’-к-3 ‘(3’ ← 5 ‘). И цепи Watson, и цепи Крика могут быть смысловыми или антисмысловыми цепями в зависимости от конкретного генного продукта, полученного из них.
Например, обозначение «YEL021W», псевдоним гена URA3, используемого в базе данных Национального центра биотехнологической информации (NCBI), означает, что этот ген находится в 21-й открытой рамке считывания (ORF) от центромеры левой руки ( L) хромосомы дрожжей (Y) с номером V (E), и что цепь, кодирующая экспрессию, представляет собой цепь Ватсона (W). «YKL074C» обозначает 74-ю ORF слева от центромеры хромосомы XI, и что кодирующая цепь является цепью Крика (C). Другой сбивающий с толку термин, относящийся к «плюсовой» и «минусовой» цепям, также широко используется. Независимо от того, является ли цепь смысловой (положительной) или антисмысловой (отрицательной), запросной последовательностью по умолчанию при выравнивании NCBI BLAST является «Плюсовая» цепь.
Амбисенс
Одноцепочечный геном, который используется как в положительном, так и в отрицательном смысле, называется амбисенсом . Некоторые вирусы имеют амбисенсный геном. Буньявирусы имеют три фрагмента одноцепочечной РНК (оцРНК), некоторые из которых содержат участки как с положительным, так и с отрицательным смыслом; аренавирусы также являются оцРНК-вирусами с амбисенсным геномом, поскольку они имеют три фрагмента, которые в основном имеют отрицательный смысл, за исключением части 5′-концов большого и малого сегментов их генома.
Антисмысловая РНК
Последовательность РНК, комплементарная транскрипту эндогенной мРНК, иногда называют « антисмысловой РНК ». Другими словами, это некодирующая цепь, комплементарная кодирующей последовательности РНК; это похоже на вирусную РНК с отрицательным смыслом. Когда мРНК образует дуплекс с комплементарной антисмысловой последовательностью РНК, трансляция блокируется. Этот процесс связан с интерференцией РНК . Клетки могут естественным образом производить антисмысловые молекулы РНК, называемые микроРНК , которые взаимодействуют с комплементарными молекулами мРНК и подавляют их экспрессию . Эта концепция также использовалась в качестве метода молекулярной биологии путем искусственного введения трансгена, кодирующего антисмысловую РНК, чтобы заблокировать экспрессию интересующего гена. Радиоактивно или флуоресцентно меченая антисмысловая РНК может использоваться для демонстрации уровня транскрипции генов в различных типах клеток.
Некоторые альтернативные антисмысловые структурные типы были экспериментально применены в качестве антисмысловой терапии . В Соединенных Штатах Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) одобрило фосфоротиоатные антисмысловые олигонуклеотиды фомивирсен (Витравен) и мипомерсен (Кинамро) для терапевтического использования у человека.
Смысл РНК в вирусах
В вирусологии термин «смысл» имеет несколько иное значение. Можно сказать, что геном РНК-вируса имеет положительный смысл , также известный как «плюс-цепь», или отрицательный смысл , также известный как «минус-цепь». В большинстве случаев термины «смысл» и «нить» используются взаимозаменяемо, делая такие термины, как «положительная цепь» эквивалентными «положительному значению», и «положительная цепь», эквивалентными «положительному значению». Независимо от того, является ли вирусный геном положительным или отрицательным, можно использовать в качестве основы для классификации вирусов.
Позитивный смысл
Положительно чувство ( 5 ‘ -До- 3’ ) вирусных РНК означает , что конкретная последовательность вирусной РНК может быть непосредственно переведены в вирусные белки (например, те , которые необходимы для репликации вируса). Следовательно, в вирусах с положительной РНК геном вирусной РНК можно рассматривать как вирусную мРНК и может немедленно транслироваться клеткой-хозяином. В отличие от РНК с отрицательным смыслом, РНК с положительным смыслом имеет тот же смысл, что и мРНК. Некоторые вирусы (например, Coronaviridae ) имеют геномы с положительным смыслом, которые могут действовать как мРНК и использоваться непосредственно для синтеза белков без помощи дополнительных промежуточных РНК. Из-за этого этим вирусам не нужно иметь РНК-полимеразу, упакованную в вирион — РНК-полимераза будет одним из первых белков, продуцируемых клеткой-хозяином, поскольку она необходима для репликации генома вируса.
Отрицательный смысл
Вирусная РНК с отрицательным смыслом (3′-к-5 ‘) комплементарна вирусной мРНК, таким образом, РНК-зависимая РНК-полимераза должна продуцировать из нее РНК-зависимую РНК-полимеразу до трансляции. Подобно ДНК, отрицательно-смысловая РНК имеет нуклеотидную последовательность, комплементарную мРНК, которую она кодирует; также как и ДНК, эта РНК не может быть напрямую транслирована в белок. Вместо этого его сначала необходимо транскрибировать в РНК с положительным смыслом, которая действует как мРНК. Некоторые вирусы (например, вирусы гриппа ) имеют негативный смысл генома и поэтому должны нести РНК-полимеразу внутри вириона.
Антисмысловые олигонуклеотиды
Молчание генов может быть достигнуто путем введения в клетки короткого «антисмыслового олигонуклеотида», комплементарного РНК-мишени. Этот эксперимент был впервые проведен Замечником и Стивенсоном в 1978 году и продолжает оставаться полезным подходом как для лабораторных экспериментов, так и, возможно, для клинического применения ( антисмысловая терапия ). Некоторые вирусы, такие как вирусы гриппа, респираторно-синцитиальный вирус (RSV) и коронавирус SARS (SARS-CoV), были нацелены на использование антисмысловых олигонуклеотидов для подавления их репликации в клетках-хозяевах.
Если антисмысловой олигонуклеотид содержит участок ДНК или имитатор ДНК (фосфоротиоатная ДНК, 2’F-ANA или другие), он может рекрутировать РНКазу H для разрушения целевой РНК. Это делает механизм сайленсинга генов каталитическим. Двухцепочечная РНК может также действовать как каталитический фермент-зависимый антисмысловой агент через путь РНКи / миРНК , включая распознавание целевой мРНК посредством спаривания смысловой и антисмысловой цепей с последующей деградацией целевой мРНК с помощью РНК-индуцированного комплекса сайленсинга (RISC). Система hok / sok плазмиды R1 представляет собой еще один пример фермент-зависимого процесса антисмысловой регуляции посредством ферментативной деградации образующегося дуплекса РНК.
Другие антисмысловые механизмы не зависят от ферментов, но включают стерическое блокирование их РНК-мишени (например, для предотвращения трансляции или для индукции альтернативного сплайсинга). В антисмысловых механизмах стерического блокирования часто используются сильно модифицированные олигонуклеотиды. Поскольку нет необходимости в распознавании РНКазы H, это может включать химические соединения, такие как 2′-O-алкил, пептидная нуклеиновая кислота (PNA), заблокированная нуклеиновая кислота (LNA) и олигомеры морфолино .
Источник


