деконденсация
Смотреть что такое «деконденсация» в других словарях:
Деконденсация стадия деконденсации — Деконденсация, стадия деконденсации * дэкандэсацыя, стадыя д. * decondensation or stage of d стадия разрыхления гетерохроматина (см.) при переходе ядра от интерфазы к профазе (см. ) … Генетика. Энциклопедический словарь
деконденсация хромосом — (де + конденсация) см. Деспирализация хромосом … Большой медицинский словарь
дифференциальная деконденсация [хромосом] — differential decondensation дифференциальная деконденсация [хромосом]. Процесс частичной деспирализации митотических хромосом во время гипотонической обработки хромосомных препаратов; предполагается, что процесс Д.д. лежит в основе проявления G… … Молекулярная биология и генетика. Толковый словарь.
дифференциальная деконденсация [хромосом] — Процесс частичной деспирализации митотических хромосом во время гипотонической обработки хромосомных препаратов; предполагается, что процесс Д.д. лежит в основе проявления G исчерченности метафазных и прометафазных хромосом. [Арефьев В.А.,… … Справочник технического переводчика
Дифференциальная деконденсация хромосом — * дыферэнцыяльная дэкандэнсацыя храмасом * differential chromosome decondensation процесс частичной деспирализации митотических хромосом во время гипотонической обработки хромосомных препаратов. Предполагается, что процесс Д. д. х. лежит в основе … Генетика. Энциклопедический словарь
деспирализация — деконденсация Процесс развертывания и раскручивания плотно упакованной молекулы ДНК в хромосоме при переходе клетки от телофазы к интерфазе; процесс Д. является обратным процессу спирализации. [Арефьев В.А., Лисовенко Л.А. Англо русский толковый… … Справочник технического переводчика
деспирализация хромосом — (син. деконденсация хромосом) процесс раскручивания спирализованных хромосом в телофазе митоза и мейоза … Большой медицинский словарь
Деспирализа́ция хромосо́м — (син, деконденсация хромосом) процесс раскручивания спирализованных хромосом в телофазе митоза и мейоза … Медицинская энциклопедия
Митоз — Фазы митоза Митоз (греч … Википедия
Мейоз — (от греч. meiosis уменьшение) или редукционное деление клетки деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с… … Википедия
Источник
Что значит хромосомы конденсируются
• При подготовке к расхождению хромосомы конденсируются и мигрируют к центру митотического веретена
• Хромосомы прикрепляются к микротрубочкам, выходящим из противоположных полюсов веретена, с помощью специальных участков, называемых кинетохорами
• Разрываются связи, соединяющие сестринские хроматиды вместе, и происходит их разделение
• Происходит расхождение изолированных сестринских хроматид, что предшествует цитокинезу
Как отмечалось выше, у большинства клеток одним из характерных признаков вступления в митоз является конденсация хромосом. Генетический материал интерфазных клеток упакован не так плотно как в митозе, и в процессе сегрегации хромосом важную роль играет его реорганизация с образованием компактных структур высшего порядка. Как достигается эта компактизация и каким образом она регулируется?
В конденсации хромосом участвует мультибелковый комплекс, который называется конденсин. Этот комплекс состоит из двух суперспирализованных белковых молекул, содержащих АТФазные домены и, в силу исторических причин, относящихся к семейству SMC-белков. Предполагается, что субъединицы конденсина могут «прогуливаться» по ДНК, скрепляя вместе ее различные участки. В состав конденсинового комплекса также входят другие белки, которые, наряду с SMC белками, вызывают в ДНК конформационные и топологические изменения, приводящие к компактизации.
Доступ конденсинового комплекса к хромосоме тщательно контролируется. Комплекс связывается с хромосомой только во время митоза. В клетках сливающихся дрожжей, у которых не разрушается ядерная оболочка, конденсин физически отделен от хромосом, будучи заключен в пределах цитоплазмы. Фосфорилирование одной из субъединиц конденсина под действием Cdk1, происходящее в митозе, вызывает миграцию комплекса в ядро. У Xenopus Cdk1, вероятно, активирует процесс суперспирализации, происходящий при действии конденсина.
О процессе конденсации хромосом также говорят и другие изменения связанных с ними белков, например фосфорилирование гистонов Н1 и Н3. Известно, что фосфорилирование гистона Н3 происходит с участием киназ Aurora. Фактически, фосфорилирование этого гистона в митозе обычно служит показателем активности киназы Aurora.
Образование связей между сестринскими хроматидами представляет собой процесс, родственный конденсации хромосом. При репликации в S-фазе каждая пара хроматид связывается между собой и находится в связанном состоянии до момента разделения при переходе от метафазы к анафазе. Каким образом и когда устанавливаются связи между хроматидами и как они разрываются в анафазе?
Так же как и конденсиновый комплекс, когезиновый комплекс формирует и поддерживает связи между сестринскими хроматидами, начиная от момента репликации ДНК и до начала их сегрегации. Этот комплекс состоит из двух SMC-белков, которые напоминают SMC-белки конденсина, хотя и отличаются от них, и трех дополнительных белков, Scc1, Scc3 и Pds5, также необходимых для его функционирования.
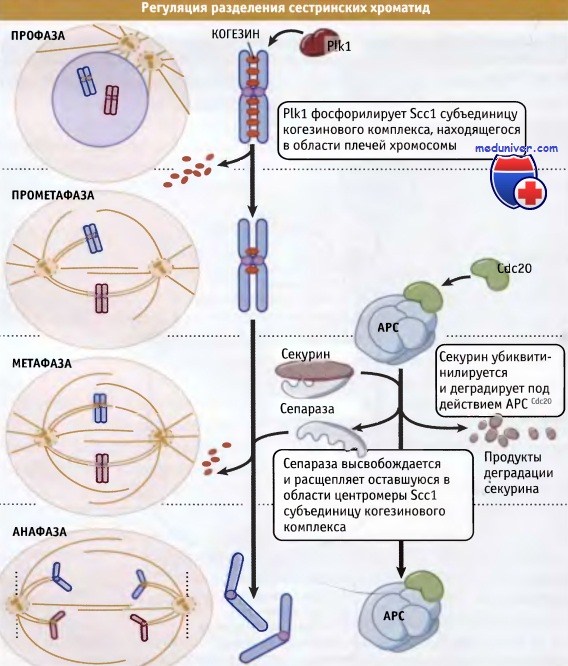 До наступления метафазы одна из протеаз, под названием сепараза, находится в связанном с секурином состоянии и не проявляет активности.
До наступления метафазы одна из протеаз, под названием сепараза, находится в связанном с секурином состоянии и не проявляет активности.
В метафазе, при действии АРС, секурин подвергается протеолизу, и таким образом происходит деблокирование сепаразы.
После этого активная сепараза расщепляет когезин, что вызывает разделение сестринских хроматид.
Предполагалось, что когезиновый комплекс скрепляет сестринские хроматиды наподобие застежки «молния». Однако недавно было показано, что SMC-белки и субъединица Scc1 обвивают сестринские хроматиды, скрепляя их. Подробности механизма этого скрепления сейчас исследуются.
Предложено два механизма разрушения связей между хроматидами. Согласно одному механизму, в профазе, с плечей хромосомы удаляется большая часть когезина, а связанный с центромерой комплекс остается на месте. Этот первый этап удаления когезина запускается Plk1, которая фосфорилирует Scc1. В то же время связанная с центромерой Scc1 когезиновая субъединица подвергается протеолизу. Если модель скрепления хроматид за счет их обвивания субъединицей Scc1 правильна, то расщепление этой субъединицы должно приводить к раскрытию обвивающего кольца и к физическому разделению сестринских хроматид. Во всяком случае, каким бы ни был механизм скрепления хроматид, очевидно, что разрушение Scc1 вызывает их диссоциацию и обеспечивает наступление анафазы.
Сепараза представляет собой сайт-специфическую протеазу, которая расщепляет Scc1 и вызывает разделение хроматид. В течение большей части клеточного цикла, сепараза поддерживается в неактивном состоянии, находясь в комплексе с другим белком, секурином. При переходе из метафазы в анафазу секурин подвергается убиквитинилированию и атакуется протеазами. При разрушении секурина высвобождается сепараза (также известная под названием сепарин), которая разрушает Scc1.
Процессы, происходящие при разделении хроматид, представлены на рисунке ниже. Таким образом, в этой простой модели ключевым процессом, запускающим сегрегацию хромосом, является протеолиз секурина. В связи с этим возникает интересный вопрос: каким образом регулируется деградация секурина?
Секурин отбирается для деградации при его взаимодействии с ЕЗ-убиквитинлигазой, известной под названием анафазный промоторный комплекс (АРС). Этот комплекс узнает белки, содержащие короткие последовательности, называемые деструктивными и/или KEN доменами (D boxes или KEN boxes). При добавлении этих последовательностей, содержащих около девяти аминокислот, в структуру белков, последние проибретают способность к деградации под действием АРС.
В отличие от SCF, для того чтобы субстрат был узнан АРС, не требуется его предварительного фосфорилирования. У почкующихся дрожжей существенная функция АРС состоит в деградации циклинов и секурина, хотя для этого комплекса известно также много других мишеней.
АРС проявляет активность только в митозе и G1-периоде, когда он предотвращает накопление циклинов и секурина. Как регулируется активность АРС? Во-первых, связывание адаптерных белков (Cdc20 и Cdh1 — обозначаемых как APC cdc20 APC cdh1 ) с APC обеспечивает значительную степень субстратной специфичности комплекса. Адаптерные белки способны связываться с АРС лишь в митозе и G1-периоде цикла. Во-вторых, активность АРС регулируется при специфическом фосфорилировании некоторых из его 13 субъединиц и адаптерных белков.
В фосфорилировании и активации АРС участвуют Cdkl и Plk1. В-третьих, активность АРС может контролироваться сигнальной системой, которая следит за прикреплением хромосом, а также точкой, контролирующей сборку веретена (SAC). Столь многочисленные уровни регуляции гарантируют, что АРС активируется только в митозе и запускает протеолиз секурина и циклина, активирует сепаразу, расщепляет когезин, и, наконец, вызывает разделение сестринских хроматид. В G1-периоде АРС сохраняет свою активность с тем, чтобы поддерживать активность Cdk1 на низком уровне. Низкая активность этой киназы позволяет сформироваться pre-RC, необходимому для следующего раунда репликации ДНК. Схема процессов активации АРС представлена на рисунке ниже.
Когда после разрыва связи сестринские хроматиды отделились друг от друга, они расходятся к противоположным полюсам клетки под действием микротрубочек веретена. Когда хромосомы разошлись и активность Cdk1 снизилась, наступает цитокинез.
Подводя итоги изложенному выше, подчеркнем, что конденсация хромосом происходит с участием белкового комплекса, который называется конденсин, и что сестринские хроматиды удерживаются вместе когезиновым комплексом до тех пор, пока не произойдет их разделение в анафазе. Разрыв связей между хроматидами требует расщепления когезина под действием протеазы, сепаразы. Для активации сепаразы необходим протеолиз ее ингибитора, секурина, который происходит с участием АРС по убиквитиновому механизму.
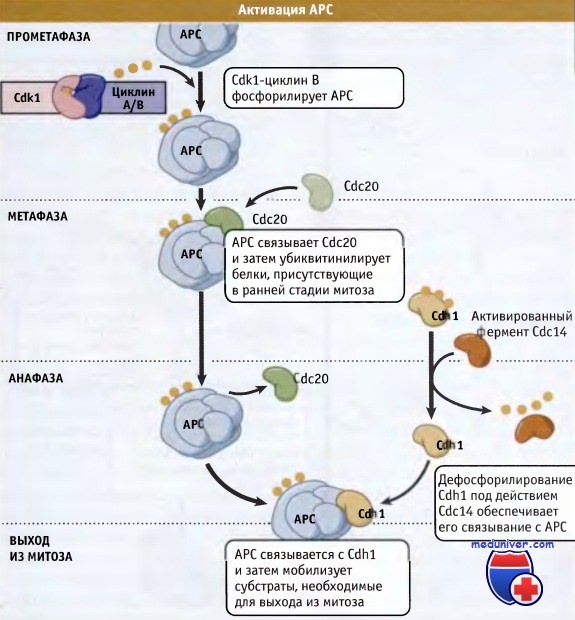 В интерфазе АРС убиквитин лигаза неактивна.
В интерфазе АРС убиквитин лигаза неактивна.
При наступлении митоза Cdk1 фосфорилирует АРС и он связывается с активатором, Cdc20.
Затем Cdc20 АРС узнает субстраты, например секурин, обеспечивая необратимое прохождение клетки по фазам митоза.
Позже в митозе АРС связывается с активатором Cdh1 и мобилизует другие субстраты, необходимые для выхода из митоза.  Первый видеокадр, показывающий хромосомы в начальных стадиях митоза.
Первый видеокадр, показывающий хромосомы в начальных стадиях митоза.
Видео процесс и фазы митоза
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Научная электронная библиотека

Юров И. Ю., Ворсанова С. Г., Воинова В. Ю., Чурносов М. И., Юров Ю. Б.,
3.4. Деление клетки
Все клетки человека проходят через цикл деления (клеточный цикл). Известны два типа деления клетки: митотческое и мейотическое, которые представлены на рисунках 6 и 7. Митоз – деление клетки, за счет которого достигается воспроизведение соматических клеток в организме. Мейоз – деление клеток, в ходе которого получаются клетки с редуцированным гаплоидным (n) набором хромосом (половые клетки). Среднее время клеточного цикла у млекопитающих составляет 17–18 часов. Клеточный цикл разделен на четыре основных стадии: G1, S (фаза синтеза ДНК), G2 и митоз (М). Первые три фазы представляют собой интерфазный период или интерфазу. Клетки, которые не делятся, находятся на стадии покоя G0.
Во время стадии G1 (примерно 9 часов) хромосомы имеют вид одиночных хроматид, клетка метаболически активна и в ней происходит синтез белков. Если клетка, находящаяся на G1 стадии, не подвергается последующему делению, то это состояние соответствует G0. Фаза синтеза (или S фаза) длится около 5 часов и характеризуется процессом репликации хромосомной ДНК. На этой стадии хромосомы состоят из двух идентичных сестринских хроматид. G2 стадия длится примерно 3 часа. Во время этой стадии клетка готовится к процессу деления. Завершение G2 соответствует концу интерфазы. Митоз длится не более 1–2 часов и является процессом образования двух генетически идентичных дочерних клеток. В свою очередь, митоз также делится на 4 стадии: профаза, метафаза, анафаза и телофаза (рис. 6).
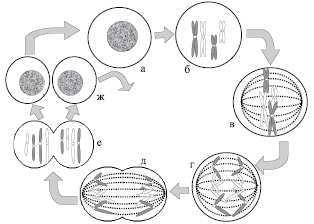
Рис. 6. Митоз. Схематическое изображение двух пар хромосом во время митоза: а – интерфаза; б – профаза; в – метафаза; г – анафаза; д – телофаза; е – цитокинез; ж – интерфазы разделившихся клеток
Профаза – стадия, во время которой происходит постепенная конденсация (уплотнение) и спирализация хромосом, в результате чего, они имеют вид дискретных структур. Во время профазы образуется веретено деления (двуполюсное веретено, состоящее из пучков микротрубочек, которые тянутся от одного полюса к другому).
Метафаза – стадия, характеризующаяся тем, что полностью осуществляется присоединение хромосом к нитям веретена, и хромосомы собираются в экваториальной плоскости клетки, находящейся на одинаковом расстоянии от обоих полюсов веретена. На этой стадии хромосомы достигают максимальной конденсации.
Цитогенетики выделяют начало метафазы как прометафазу, на которой довольно часто проводят лабораторные исследования. Прометафаза – короткий промежуточный период между профазой и метафазой, в ходе которой исчезает ядерная мембрана и появляются нити веретена деления, к которым с помощью кинетохоров присоединяются хромосомы.
Анафаза – стадия разделения сестринских хроматид и их расхождения к противоположным полюсам веретена.
Телофаза – последняя стадия митоза. Она начинается, когда все сестринские хроматиды доходят до полюсов веретена. В ходе данной фазы митоза происходит восстановление клеточного ядра и внутриядерных структур. За ней, как правило, следует цитокинез – разделение двуядерной клетки на две с одним ядром в каждой (рис. 6).
Мейоз – деление клеток, в ходе которого получаются клетки с редуцированным гаплоидным набором хромосом (половые клетки). Данный процесс включает в себя две фазы клеточного деления: мейоз 1 и мейоз 2. В ходе мейоза происходит редукция диплоидного хромосомного набора (2n) до гаплоидного (n).
Мейоз 1, в свою очередь также делится на несколько стадий: профаза I, метафаза I, анафаза I и телофаза I (рис. 7). Профаза I является комплексной стадией, которая, в свою очередь, делится на несколько этапов:
лептотена: 46 хромосом состоят из 2-х хроматид и начинают коденсироваться; эта стадия характеризует начало мейоза;
зиготена: гомологичные хромосомы спариваются своими участками – этот этап деления называется синапсом; в результате образуется тройственная структура – синаптонемальный комплекс;
пахитена: стадия завершения синапса, спаренные гомологи, состоящие из 4-х хроматид, формируют бивалент; во время этой стадии происходит кроссинговер – обмен последовательностями ДНК хромосом между несестринскими хроматидами бивалентов. Результатом этого процесса является рекомбинация генетического материала между гомологичными хромосомами, создающая новые комбинации генов в дочерних клетках;
диплотена: на этой стадии хромосомы отталкиваются друг от друга до тех пор, пока гомологи не будут соединены только участками, подверженными кроссинговеру. Такие участки называются хиазмами;
диакинез: хромосомы претерпевают наибольшее сжатие во время этой последней стадии профазы I.
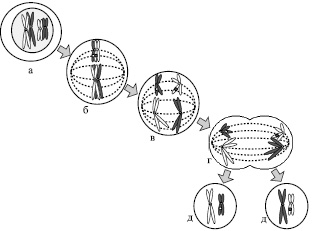
Рис. 7. Мейоз I. Схематическое изображение двух пар хромосом во время мейоза I: а – профаза I; б – метафаза I; в – анафаза I; г – телофаза I; д – клетки, образующиеся в результате первого мейоза
Метафаза I характеризуется исчезновением ядерной мембраны и образованием мейотического веретена деления. Биваленты выравниваются по экваториальной плоскости клетки и их центромеры случайным образом ориентируются к противоположным полюсам. Во время анафазы I биваленты разделяются и расходятся к противоположным полюсам. В ходе телофазы I каждая хромосома из двух гаплоидных наборов достигает противоположных полюсов, и образуются две дочерние клетки, в каждой из которых по 23 хромосомы, состоящие из 2-х хроматид.
Мейоз 2 практически идентичен митотическому делению за исключением того, что в данном случае делящиеся клетки имеют гаплоидный хромосомный набор. Хромосомы выравниваются по экваториальной плоскости клетки на стадии метафазы II, хроматиды разделяются и расходятся к противоположным полюсам на стадии анафазы II, цитокинез происходит на стадии телофазы II. В результате митотического деления (мейоза 1 и 2), как правило, образуются 4 дочерние клетки с гаплоидным набором хромосом, каждая из которых генетически отличается друг от друга за счет процесса кроссинговера и случайного расхождения гомологичных хромосом.
Источник


